Způsob prodloužení a ukončení translace mRNA během syntézy proteinů
Krok translace mRNA během prodloužení. Po zahájení polypeptid je sestaven ve formě po sobě následujících aminokyselin, která je určena sekvencí mRNA. Tento proces, ve kterém jsou každé připojené aminokyseliny rozděleny dvě GTP molekuly vyžaduje značné množství energie.
stadium prodloužení translace mRNA To zahrnuje minimální počet proteinových faktorů, než zahajovací fáze (tři EEF vs více než deset elF), ale tyto jsou považovány za nezbytné EEF „dříč“ syntézy proteinu na ribozomu. Na začátku prodloužení methionyl-tRNA párových spojen se start kodonem mRNA k ribosomu nazývá peptidyl nebo části P. Potom protein faktor eEFIA (spojené s GTP) dodává správné další aminoacyl-tRNA na ribozomy, aminoacyl oblasti nebo části A.
Po správném kodonově antikodon interakce eEFIA energie uvolněná hydrolýzou GTP.
Druhým faktorem, nazvaný eEFIB, Pomáhá při regeneraci aktivního eEFIA poskytuje nepřetržitou dodávku aminoacyl-tRNA na ribozomu.
Třetí (a poslední) faktor, nazvaný eEF2, zprostředkovává translokaci peptidyl-tRNA z místa P k místu A a usnadňuje pohyb ribozomu podél mRNA. Pro tento ribosomální translokaci také potřebují energii pocházející z hydrolýzy GTP.

úroveň eEF2 v buňce odráží míru syntézy proteinu aktivity. V podstatě, v mléčné žláze obsahoval 20 krát více eEF2, než v játrech, a spekuluje se, že eEF2 je omezujícím faktorem v syntéze mléčných proteinů.
Činnost všech tří eef v savčích buňkách vystaveny regulaci fosforylací. Fosforylace eEFIA inzulín stimuluje aktivitu prodloužení, ale neexistují žádné zprávy, že fosforylace eEFIB ovlivňuje rychlost protažení, ne. Na druhou stranu, inzulin způsobuje defosforylace eEF2, stimulovat aktivitu prodloužení.
fosforylace eEF2 je nejvíce studoval a pravděpodobně nejdůležitější mechanismus pro snížení prodloužení sazby. EEF2 fosforylace je katalyzována specifickou Ca2 + -kalmodulin EEF2 závislé-kinázy. Aktivita eEF2 regulované kinázy mTOR-kináza, ale jak je to mTOR defosforylace zůstává nejasný.
Krok ukončení translace mRNA
Poslední etapa vysílání - ukončení. Začíná, když je stop kodon umístěný v oddíle A ribozomu. Tato událost je rozpoznán proteinu eRF1, který se váže na ribozom a GTP způsobem závislým katalyzuje štěpení vazby mezi peptidy a tRNA, čímž se uvolní protein.
Druhý se nazývá faktor uvolňující ERF-3, slouží k podpoře aktivity eRF1 v přítomnosti GTP.

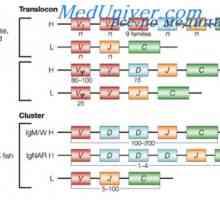 Translokon imunoglobuliny. Kombinace v- a geny protilátek
Translokon imunoglobuliny. Kombinace v- a geny protilátek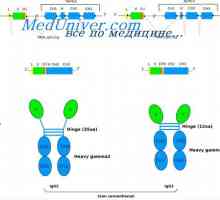 Syntéza Poliribosomny komplex protilátka. RNA podílí na syntéze protilátky
Syntéza Poliribosomny komplex protilátka. RNA podílí na syntéze protilátky Tvorba imunoglobulinových řetězcích. Přebytek protilátky syntéza L-řetězce
Tvorba imunoglobulinových řetězcích. Přebytek protilátky syntéza L-řetězce Syntéza imunoglobulinu těžkého a lehkého řetězce. Jednotná syntéza těžkých a lehkých řetězců…
Syntéza imunoglobulinu těžkého a lehkého řetězce. Jednotná syntéza těžkých a lehkých řetězců… MRNA se podílí na syntéze protilátky. Metody studia mRNA
MRNA se podílí na syntéze protilátky. Metody studia mRNA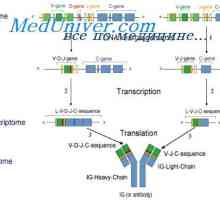 Vlastnosti protilátkou mRNA. MRNA struktura imunoglobulinů
Vlastnosti protilátkou mRNA. MRNA struktura imunoglobulinů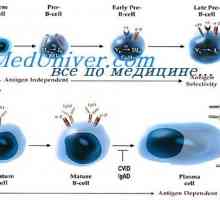 Trvání mRNA protilátky. Vliv na syntézu mRNA imunoglobulinu
Trvání mRNA protilátky. Vliv na syntézu mRNA imunoglobulinu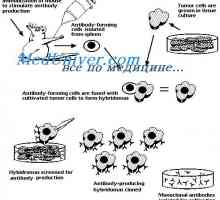 Překlad mRNA. Vlastnosti translace mRNA syntetizované protilátky
Překlad mRNA. Vlastnosti translace mRNA syntetizované protilátky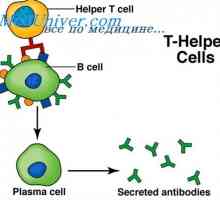 Prekurzory protilátek syntézy L-řetězec. Ekstrapeptidy a jejich funkce
Prekurzory protilátek syntézy L-řetězec. Ekstrapeptidy a jejich funkce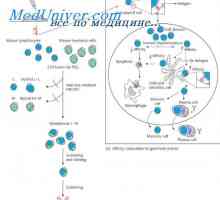 Geny syntézu protilátky. Počet genů, které se účastní syntézy imunoglobulinů
Geny syntézu protilátky. Počet genů, které se účastní syntézy imunoglobulinů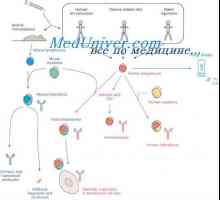 Hybridizace mRNA a DNA protilátek. Lokalizace v- a geny v genomu imunoglobulinů
Hybridizace mRNA a DNA protilátek. Lokalizace v- a geny v genomu imunoglobulinů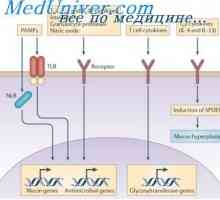 Simultánní syntéza různých imunoglobulinů. Krvetvorné kmenové buňky
Simultánní syntéza různých imunoglobulinů. Krvetvorné kmenové buňky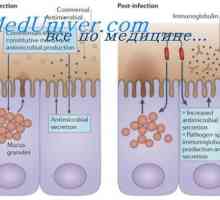 Struktura membránových imunoglobulinů. Původ povrchových protilátek
Struktura membránových imunoglobulinů. Původ povrchových protilátek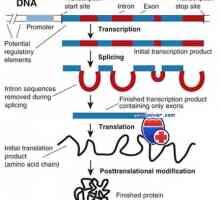 Transkripce. Formy a druhy RNA buněk
Transkripce. Formy a druhy RNA buněk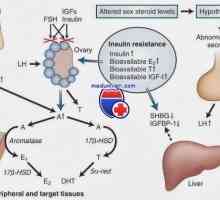 Vaječníků růstové faktory. Inzulín a epidermální růstový faktor
Vaječníků růstové faktory. Inzulín a epidermální růstový faktor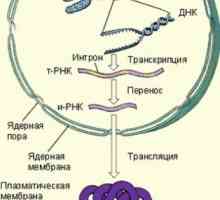 Kroky syntézy bílkovin v těle
Kroky syntézy bílkovin v těle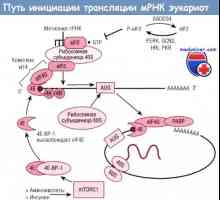 Cesta iniciace translace mRNA během syntézy proteinů
Cesta iniciace translace mRNA během syntézy proteinů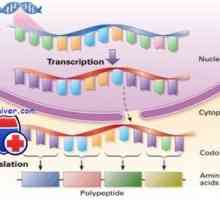 Regulace transkripce a translace v oocytu
Regulace transkripce a translace v oocytu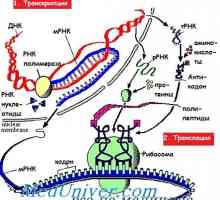 Etapy translace mRNA během syntézy proteinů
Etapy translace mRNA během syntézy proteinů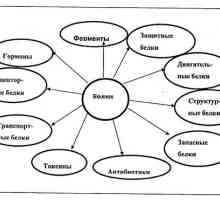 Hlavními chemickými složkami živých organismů. různé faktory
Hlavními chemickými složkami živých organismů. různé faktory Změna funkcí chromatinu v průběhu stárnutí. změna překlad
Změna funkcí chromatinu v průběhu stárnutí. změna překlad