Cesta iniciace translace mRNA během syntézy proteinů
Většina řídících faktorů překlad To se odkazuje na zahajovací fázi, která pak může být rozdělena do tří fází pro všechny činnosti zahajovací fázi. V první fázi je v důsledku vazby na iniciační tRNA (Zejména methionyl-tRNA) a malé ribosomální podjednotky. Methionyl-tRNA je dodáván s ribozomální 40S podjednotku proteinového faktoru eIF2.
kapacita eIF2 a guanosin-5`-trifosfát (GTP) Se vztahují k zahájení methionyl-tRNA regulované stav fosforylace eIF2. Zisk fosforylovaný eIF2 zabraňuje překrývání methionyl-tRNA na malou ribozomální podjednotky, což vede k celkové inhibici translace mRNA. V podmínkách buněčný stres fosforylace eIF2 také ukazuje, že je třeba zvýšit expresi specifických mRNA, které pracují pro regulaci a zmírnění buněčný stres.
Tyto procesy jsou složité reakce na stres a spouští rodiny čtyř eIF2 kinas: Perk (PKR-podobný protein kináza endoplazmatického retikula, také nazýván pankreatu eIF2 kináza, OBS), který detekuje stresu endoplazmatického retikula (ER) - GCN2 (obecný řídící nonderepressible-2 kinázu), který se aktivuje aminoskupinu nevýhodu a UV izlucheniem- hem kontrolované inhibitor, který detekuje ztráta motiv je PKR (dsRNA-dependentní proteinkináza), který je aktivován pomocí virové infekce.
tito proteinkináza funkce především pro detekci specifického stresor na životní prostředí, ale bylo zjištěno, že tyto proteiny se jednat společně a jsou schopny provádět podpůrnou kinázovou aktivitu, když je provoz primárního reaktivní kinázy rozbité nebo chybí.
Druhá etapa v iniciaci překlad, který podléhá regulaci, spočívá v připojení k malé ribozomální podjednotky vybrané mRNA. To vyžaduje komplexní vícesložkový, který má obecný název eIF4 (nebo eIF4F). Jeden z proteinů v této skupině, uvedené eIF4E, vybere mRNA být přeloženy svou vazbou 5`-uzavírací struktury. Všechny eukaryotické mRNA mají cap strukturu 7-methylguanosine.

Po vytvoření komplexu před zahájením plánované pro vysílání skupiny vybrány translace mRNA faktory elF4, který obsahuje mRNA proteinu kepsvyazyvayuschy, elF4E a „rám“ proteinu, elF4C. elF4C hraje důležitou roli v přibližné malých ribozomální podjednotky a mRNA prostřednictvím interakce s elF4E a elF3, a v cirkularizaci mRNA vazbou na poly (A) vážící protein (PABP).
Živiny závislé komplex 1 mTOR (mTORCl) reguluje aktivitu tohoto kroku části komunikačním regulace elF4C elF4E s ohledem na překlad represoru 4E-BP-1. Po úspěšné vytvoření aktivního komplexu elF4 a výběrem mRNA ribozomální podjednotky 60S podjednotku 40S se připojí k formě příslušné ribozómu 80S. HRI - hem kontrolované inhibiční PERK - PKR-jako endoplazmatického retikuluma- proteinkinázu PKR - dsRNA-dependentní protein kinázy.
kovalentně připojen molekuly 7-methylguanosine To slouží k ochraně proti mRNA exonukleasami, ale co je důležitější, 7-methylguanosine rozpoznat mRNA kepsvyazyvayuschim protein (eIF4E), pro provedení výběru a vázání malé ribozomální podjednotky. Druhý člen eIF4 skupinu s názvem eIF4G, postupujících malé ribozomální podjednotky mRNA. Dosahuje toho tím, že působí jako „kostry» (lešení), která se skládá z 13 podjednotek komplexu proteinů, které se vážou eIF4E, eIF4A a eIF3, čímž se usnadňuje jejich spojení s 40S ribozomální. Rodina represorový protein (4E-TK, zejména 4E-BP-1) může zabránit interakci mezi eIF4E a eIF4G čímž inhibují vazbu 40S ribozomální a mRNA.
Druhá funkce eIF4G je sloučenina s poly (A) vážící protein, tj. protein, který se váže na oblast zbytků 20-250 adenosinu na 3`-konci mRNA. Tyto dvě související jevy jsou dokončeny 5` => 3`-cirkularizaci mRNA během překladu. Předpokládá se, že cirkularizaci důležitá pro stabilizaci mRNA přijímáni ribozomální podjednotky 40S, jakož i pro efektivní využití ribozomy při ukončení kroku pro další vysílání cyklu s použitím stejné mRNA. Tak eIF4G interakce s poly (A), vázající protein se podílí na zvýšení tvorby komplexů zahájených 40S a 80S a recykluje oběhu ribozomu mRNA.
Závěrečná fáze stupeň iniciace translace To zahrnuje sdružování malou ribozomální podjednotky (související mRNA) z velké ribozomální podjednotky. Tato akce je katalyzován několika EIF včetně eIF2, eIF3 a eIF5. Tyto faktory a jejich interakce byly studovány, ale podrobnosti jsou přímo spojené s možnou regulaci v této fázi překladu, nad rámec této publikace.
Pokud jde o krmení novorozenci je třeba poznamenat, že cílem stimulovat syntézu proteinu na úrovni iniciace translace potřeba inzulínu i aminokyseliny. Aminokyseliny, zejména leucin (aminokyseliny s rozvětveným řetězcem), slouží jako signální molekuly, které regulují translace mRNA počáteční kroky směrování eIF4 tvorbu komplexů a fosforylaci eIF2.
Jediná věc, která je třeba stimulace eIF4 a syntézy proteinu v kosterních svalech z nově narozených selat, - fyziologického zvýšení leucin. Tento závěr je pro organizaci novorozenecké výživy velmi důležité, protože to znamená, že aminokyseliny v buňce mají extrémně vysokou hodnotu pro stimulaci růstu tkání vyčerpány. Přesně tak, jak aminokyseliny zahájit signál pro aktivaci translace stroje, stále zůstává záhadou. Je známo, že mTOR kinázy, je jedním z hlavních členů, které detekují přítomnost aminokyselin v buňce, a že stimulace syntézy proteinů poskytnutím novorozence nebo mladých zvířat leucin optimálně aktivuje, pokud aminokyselinové signál kyseliny v kombinaci s aktivací transdukční dráhy signálu insulinu.
Video: Proton spád a syntéza ATP
Signální dráhy aktivované inzulín a aminokyseliny, mTOR sbíhají na úrovni, která pak katalyzuje fosforylaci 4E-BP-1. Fosforylace spustí uvolňování 4E-BP-1 s eIF4E, což umožňuje eIF4E a eIF4G vázat eIF3, tvořit komplexní eIF4F. Tyto vazebné procesy jsou částečně nebo úplně blokována působením rapamycin, inhibitorem mTOR. Stimulace aktivity mTOR vede k aktivaci fosforylace S6K1, který aktivuje biogeneze ribozomů a dalších procesů, které mohou vést ke zvýšení velikosti buněk. Tato signální dráhy S6K1, reagující na aminokyselinu nebo inzulinu rapamycin zcela inhibována zatímco vazba eIF4G aktivita je potlačena v menší míře.
Je zřejmé, že novorozenec tělo od okamžiku narození je připraven reagovat na toku živin aktivní a efektivní syntézy Protein. Nicméně složky překladu a faktory nezbytné pro růst aktivaci anabolických procesů, oslabují s věkem. Toto snížení translační kapacity v reakci na krmení u novorozenců je velmi rychle (v novorozených selat - během několika dnů). Tyto údaje potvrzují pozorování zásadní význam časné výživy bílkovin, které poskytují skutečnou příležitost maximálně stimulovat vývoj organismu dětí narozených s nízkou porodní hmotností.
Snad právě proto, že o této skutečnosti Nedostatek bílkovin v kojenecké stravy v raném stádiu života, je obtížné překonat v budoucnu, a to i když je vysoce kalorií, a hormonální substituční terapie nejsou schopny eliminovat zpoždění růstu, protože schopnost těla reagovat na kvalitu potravin v pozdějším životě výrazně snížena.

 Receptory pro hormony spojené s G-proteinem. Enzym-konjugovaná receptory hormonů
Receptory pro hormony spojené s G-proteinem. Enzym-konjugovaná receptory hormonů Syntéza Poliribosomny komplex protilátka. RNA podílí na syntéze protilátky
Syntéza Poliribosomny komplex protilátka. RNA podílí na syntéze protilátky Tvorba imunoglobulinových řetězcích. Přebytek protilátky syntéza L-řetězce
Tvorba imunoglobulinových řetězcích. Přebytek protilátky syntéza L-řetězce Syntéza imunoglobulinu těžkého a lehkého řetězce. Jednotná syntéza těžkých a lehkých řetězců…
Syntéza imunoglobulinu těžkého a lehkého řetězce. Jednotná syntéza těžkých a lehkých řetězců… MRNA se podílí na syntéze protilátky. Metody studia mRNA
MRNA se podílí na syntéze protilátky. Metody studia mRNA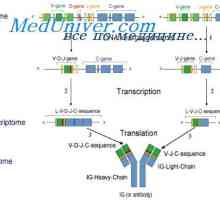 Vlastnosti protilátkou mRNA. MRNA struktura imunoglobulinů
Vlastnosti protilátkou mRNA. MRNA struktura imunoglobulinů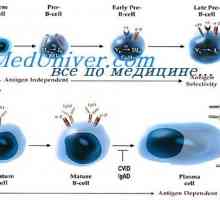 Trvání mRNA protilátky. Vliv na syntézu mRNA imunoglobulinu
Trvání mRNA protilátky. Vliv na syntézu mRNA imunoglobulinu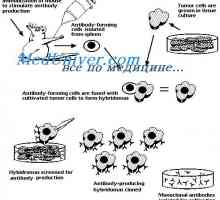 Překlad mRNA. Vlastnosti translace mRNA syntetizované protilátky
Překlad mRNA. Vlastnosti translace mRNA syntetizované protilátky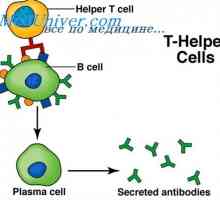 Prekurzory protilátek syntézy L-řetězec. Ekstrapeptidy a jejich funkce
Prekurzory protilátek syntézy L-řetězec. Ekstrapeptidy a jejich funkce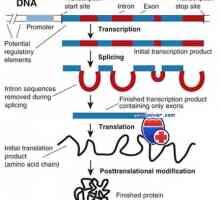 Transkripce. Formy a druhy RNA buněk
Transkripce. Formy a druhy RNA buněk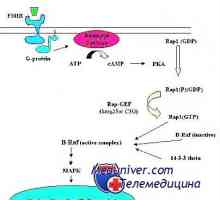 Mechanismus účinku gonadotropinu receptorů. vyčerpání receptory
Mechanismus účinku gonadotropinu receptorů. vyčerpání receptory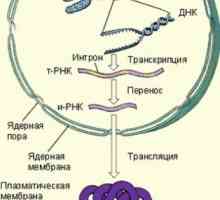 Kroky syntézy bílkovin v těle
Kroky syntézy bílkovin v těle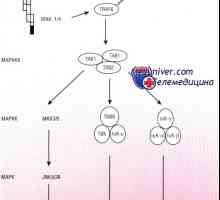 Schéma MAPK cesty v reakci na bakterie střevního epitelu
Schéma MAPK cesty v reakci na bakterie střevního epitelu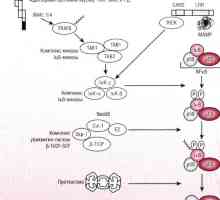 Patogeneze způsob NFKB / rel v reakci na bakterie střevního epitelu
Patogeneze způsob NFKB / rel v reakci na bakterie střevního epitelu Způsob prodloužení a ukončení translace mRNA během syntézy proteinů
Způsob prodloužení a ukončení translace mRNA během syntézy proteinů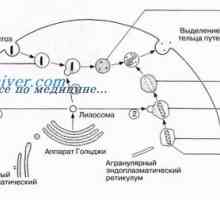 Autofagalno-lysozomální systém štěpení proteinů apoptóza
Autofagalno-lysozomální systém štěpení proteinů apoptóza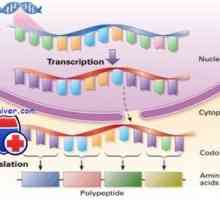 Regulace transkripce a translace v oocytu
Regulace transkripce a translace v oocytu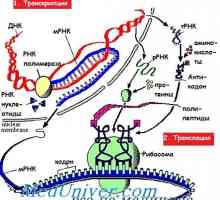 Etapy translace mRNA během syntézy proteinů
Etapy translace mRNA během syntézy proteinů Lék na vysoký krevní tlak pomáhá při roztroušené skleróze
Lék na vysoký krevní tlak pomáhá při roztroušené skleróze- Změna funkcí chromatinu v průběhu stárnutí. změna překlad
- Věkové změny v genetickém aparátu buněk. změny chromozomů