Tvorba imunoglobulinových řetězcích. Přebytek protilátky syntéza L-řetězce
Zajímavé bylo určit předpokládanou dobu syntézy a uvolňování těžké a lehké řetězce s polyribosomes. Pro tento buněk (myelom MRSI) byly označeny po dobu 90 sekund za použití směsi značených aminokyselin, se potom umístí do média, obsahujícího 200-násobného přebytku neznačeného aminokyselin, byly izolovány v různých časových intervalech těžkých a lehkých polyribosomes a množství zbývajícího na polyribosomes štítku a transformován do média ,
Obrázek ukazuje, že s plicní polyribosomes značkou "příchod„Asi dvakrát častěji než ty těžké. Odpovídající doba syntézy byla stanovena na 30-45 a 60-75 sekund. Tak, při tvorbě jednoho těžkého řetězce mohou tvořit dvě plíce, t. E. rozdíly v rychlosti syntézy se zdá být vést k nadměrné tvorbě lehkých řetězců v buňce. Je však třeba mít na paměti, že polyribosomes podílející se na syntéze H-řetězce, asi dvakrát polyribosomes, které jsou syntetizovány L-řetězec.
toto podle pořadí,, To může vést k tomu, že po krátkém počátečním období nevyvážené syntézy H- a L řetězce nastane stav, ve kterém je počet za jednotku času z syntetizovaných lehkých řetězců se rovnal počtu těžké.
Přebytek syntéza L-řetězce a jejich vstup do extracelulárního prostředí byla zjištěna v řadě studií prováděných na lymfatické uzliny a sleziny z králíků (Nezlin, Kulpina, 1966- Shapiro ea, 1966b- Skvortsov, Gurvich, 1968) a myší myelomové buňky, některé (Schubert, 1968- Baumal, Scharff, 1973). Na druhé straně, v experimentech s myelomovými buňkami a H5563 myší lymfatické Askonas a Williamson (Askonas, Williamson, 1967a) nenašel významný nadbytek volných lehkých řetězců a jejich sekreci z buňky.

V řadě případů show, že přebytek L-řetězce jsou zničeny v buňkách (Schubert, 1968- Baumal, Scharff, 1973) - popisuje zničení H řetězců na abnormální syntéze variantních buněk myelomu klonovat pouze těžkých řetězců (Baumal, Scharff, 1976).
zjištění s velkým zájmem, protože umožňují zvýšit otázky týkající se životnosti polyribosomes a mRNA různých velikostí, počet matric na H- a L-řetězce a kolikrát může sloužit jako templát je stejné mRNA, počet genů kódujících odpovídající mRNA, rozdíly v jejich rychlost transkripce a translace, účinnosti vazby různých mRNA na ribozomy, a konečně, existence rozdílům mezi jednotlivými druhy životnosti mRNA u zvířat.
v buňkách normální lymfoidní tkáně a buňky myelomu (jako ve všech buňkách), existují dvě třídy polyribosomes - volné a vázané na membrány z endoplazmatického retikula (mikrosomy). Detekce imunoglobulinu polypeptidových řetězců v nádržích endoplazmatické retikulum a frakcionační datové buňky (Swenson, Kern, 1967a), ukázaly, že nově syntetizované téměř výlučně imunoglobuliny jsou spojeny s membránou, zejména hrubé. Přímý důkaz nebylo získáno ze tří typů pokusů.
V první řadě to svědčil Data pro studium syntézy imunoglobulinů v „přirozených“ bezbuněčných systémech, tj. Např. Systémy odvozené z buněk produkujících imunoglobuliny.
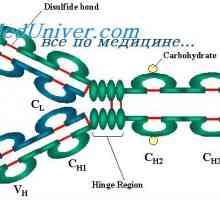 Imunoglobulin lehké řetězce. Organizace imunoglobuliny
Imunoglobulin lehké řetězce. Organizace imunoglobuliny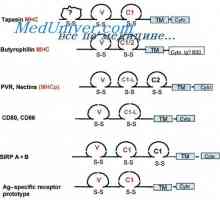 Řízení vzdělávání imunoglobulinových lehkých řetězců. Geny světlo řetězce protilátky
Řízení vzdělávání imunoglobulinových lehkých řetězců. Geny světlo řetězce protilátky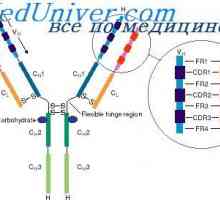 Lokalizace genů světlo řetězce protilátky. Důvody pro proměnlivosti lehkých řetězců
Lokalizace genů světlo řetězce protilátky. Důvody pro proměnlivosti lehkých řetězců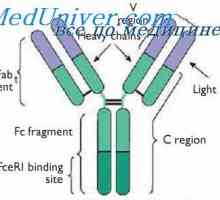 Variabilní oblasti protilátek lehkých řetězců. Geny pro variabilní oblast
Variabilní oblasti protilátek lehkých řetězců. Geny pro variabilní oblast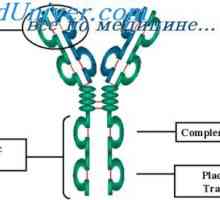 Cast lehké imunoglobulinové řetězce. Ekstrauchastki protilátky
Cast lehké imunoglobulinové řetězce. Ekstrauchastki protilátky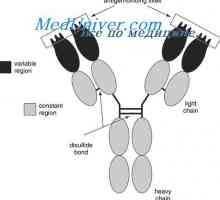 Tvorba těžkého řetězce imunoglobulinu. Konstantní oblast těžkého řetězce protilátky
Tvorba těžkého řetězce imunoglobulinu. Konstantní oblast těžkého řetězce protilátky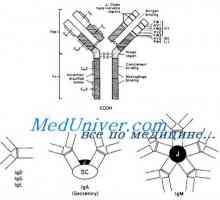 Zapletení Geny protilátky těžkého řetězce. Geny těžkých řetězců IgA
Zapletení Geny protilátky těžkého řetězce. Geny těžkých řetězců IgA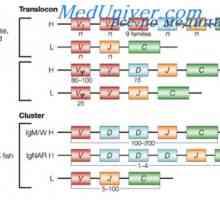 Translokon imunoglobuliny. Kombinace v- a geny protilátek
Translokon imunoglobuliny. Kombinace v- a geny protilátek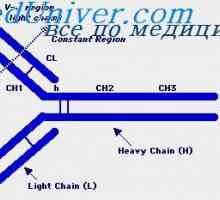 Upevňovací VH geny imunoglobulinu. Chromozom s geny protilátek
Upevňovací VH geny imunoglobulinu. Chromozom s geny protilátek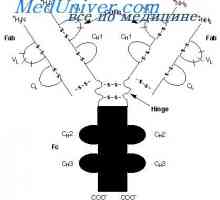 Rozdíly Variabilní oblasti protilátek. Variabilní imunoglobulinu podskupiny
Rozdíly Variabilní oblasti protilátek. Variabilní imunoglobulinu podskupiny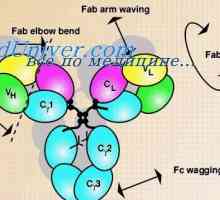 Variabilní oblasti těžkého řetězce. Variabilní oblastních genů těžkých řetězců
Variabilní oblasti těžkého řetězce. Variabilní oblastních genů těžkých řetězců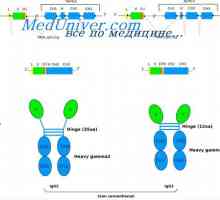 Syntéza Poliribosomny komplex protilátka. RNA podílí na syntéze protilátky
Syntéza Poliribosomny komplex protilátka. RNA podílí na syntéze protilátky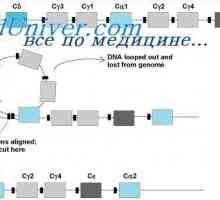 Metody pro izolaci polyribosomes. Rozměry polyribosomes syntetizovat protilátky
Metody pro izolaci polyribosomes. Rozměry polyribosomes syntetizovat protilátky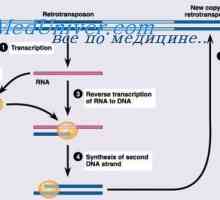 Syntéza imunoglobulinu těžkého a lehkého řetězce. Jednotná syntéza těžkých a lehkých řetězců…
Syntéza imunoglobulinu těžkého a lehkého řetězce. Jednotná syntéza těžkých a lehkých řetězců… MRNA se podílí na syntéze protilátky. Metody studia mRNA
MRNA se podílí na syntéze protilátky. Metody studia mRNA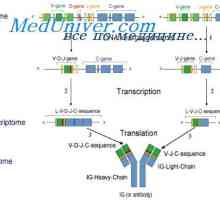 Vlastnosti protilátkou mRNA. MRNA struktura imunoglobulinů
Vlastnosti protilátkou mRNA. MRNA struktura imunoglobulinů Plazmové myší nádory. role Polyribosomes v biosyntéze imunoglobulinů
Plazmové myší nádory. role Polyribosomes v biosyntéze imunoglobulinů Prekurzory protilátek syntézy L-řetězec. Ekstrapeptidy a jejich funkce
Prekurzory protilátek syntézy L-řetězec. Ekstrapeptidy a jejich funkce Hybridizace mRNA a DNA protilátek. Lokalizace v- a geny v genomu imunoglobulinů
Hybridizace mRNA a DNA protilátek. Lokalizace v- a geny v genomu imunoglobulinů Sestavení protilátky. doplnění imunoglobulinů
Sestavení protilátky. doplnění imunoglobulinů Struktura membránových imunoglobulinů. Původ povrchových protilátek
Struktura membránových imunoglobulinů. Původ povrchových protilátek