Geny syntézu protilátky. Počet genů, které se účastní syntézy imunoglobulinů
primární výzkum Struktura a genetika imunoglobulinů To vedlo k závěru, že každý nolipeptidnaya řetězec je kódován alespoň dva geny: V-genu odpovědného za variabilní oblast a C-gepami odpovědné za konstantních oblastí molekul. Počet obou genů v genomu buněk je zásadní zájmu. Skutečnost, že pozorovaný diverzita protilátek různých autorů vysvětleno buď v podmínkách „zárodečné“ teorie genů, podle které se v genomu sady genů pro všech známých V-domény, tj. E. mnoho tisíc V-genu, a to buď z hlediska somatické teorie mutace přičemž sada v-geny v genomu omezen, a paleta struktur imunoglobuliny (protilátky), se vyskytuje jako výsledek somatické mutace. K dispozici je také kompromis hledisko. Všechny tyto myšlenky vyčerpávajícím diskutovaných, aby se stal vůdcem a spoluautory (Leder e. A., 1974), takže čím více detail nebudeme zastavit.
Všimněte si, že AE Gurvitch a R. S. Nezlin (1965) byla provedena původní předpoklad že jednotlivé úseky genomu kódují protilátky a aktivních center a že L-II- a TY „shromáždit“ menších, nezávisle syntetizované řetězce. Je velmi zajímavé, že po pěti letech, tyto myšlenky znovu objevila v podobě hypotézy „provádění» ( «vložení») informace kódované polynukleotidy podle malého (asi 30 až 45 nukleotidů), zajištění dostupnosti CDR v H a L řetězců (Wu, Kabát, 1970).
závěrečný Odpověď na tyto otázky, Je zřejmé, že může poskytovat pouze přímé stanovení V- a C-geny v genomu. Izolace mRNA přípravků s vysokým stupněm čistoty odpovídajících povolena počáteční experimenty. Přístup vyvinutý mnoho let, je určení počtu genů RNA-DNA hybridizace. Vzhledem k tomu, struktura mRNA transkribované strukturu zcela komplementární DNA část, pak může být tato metoda určení počtu genů, a na zjištění, zda jejich zesílení dochází (množení) pro indukci syntézy imunoglobulinů.

V současné době je výzkum Tento druh se provádí s mRNA, kódující syntézu L-řetězce jsou dostatečně čisté přípravky N-mRNA se ještě získá. Nezbytnou podmínkou pro provádění těchto pokusů je, za prvé, čistota přípravu mRNA a vysokou specifickou aktivitu, a za druhé, přítomnost velkého přebytku DNA. Získat vysokomechenye mRNA drog je obtížné. Typicky je to provedeno za použití mRNA izolované NT buněk kultivovaných v přítomnosti vyšších dávkách nebo 32P-3H predshsstvennikov nebo více - mRNA značených 125I in vitro (Farace ea, Matthyssens 1976- e a, 1976 ....). K dispozici je také nepřímá verze metody. To spočívá v tom, že nejprve pomocí reverzní transkriptázy na mRNA připravené matrice vysokomechenuyu komplementární DNA (cDNA), a to je již použit pro hybridizaci na buněčné DNA (Schechter např. A., 1976- Storb např. A., 1976).
Přečištěný mRNA nebo cDNA hybridizovány denaturovaných určitých podmínek (působení ultrazvuku) nebo DNA z jater myšího myelomu NS buněk a procento vázané RNA (nebo cDNA), jako funkce času hybridizace. (Stanovení je založeno na stabilitu hybridů RNase efektu.) Za těchto podmínek bylo procento hybridizované RNA (cDNA) (f) je závislá na koncentraci RNA (cDNA) (RO), koncentrace DNA (C) a inkubační době (F). S velmi velký přebytek DNA nezávisí na f R0, a závisí pouze na C „a t (Cot). Lůžku množství, při kterém hybridizace je 50%, samozřejmě závisí na stupni komplementarity DNA a mRNA (cDNA).
stupeň homologie dvou mRNA (A tedy i jejich V- nebo S-gen) se stanoví v kompetitivní inhibiční experimenty hybridizace značeného přípravu mRNA neznačeného léků homologní a heterologní mRNA, nebo izolací DNA z vytvořeného hybridu, a jeho hybridizace s jinými značeným homologní a heterologní mRNA. Porovnání těchto výsledků s údaji o primární struktury těchto polypeptidů, které jsou kódovány mRNA zkoumána, je možno vidět, zda je křivka definována frekvence opakování hybridizace genů s odhadem z údajů o homologie V- a C-domény.
 Abnormality SOx genů a TVH Holt-Oram syndromu. Fibroblastové růstové faktory
Abnormality SOx genů a TVH Holt-Oram syndromu. Fibroblastové růstové faktory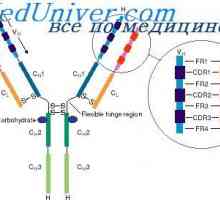 Lokalizace genů světlo řetězce protilátky. Důvody pro proměnlivosti lehkých řetězců
Lokalizace genů světlo řetězce protilátky. Důvody pro proměnlivosti lehkých řetězců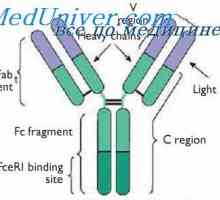 Variabilní oblasti protilátek lehkých řetězců. Geny pro variabilní oblast
Variabilní oblasti protilátek lehkých řetězců. Geny pro variabilní oblast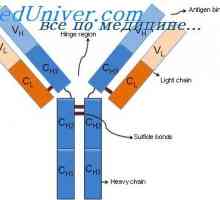 Alelické varianty imunoglobulin G. Geny těžkých řetězců IgG
Alelické varianty imunoglobulin G. Geny těžkých řetězců IgG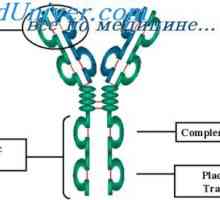 Cast lehké imunoglobulinové řetězce. Ekstrauchastki protilátky
Cast lehké imunoglobulinové řetězce. Ekstrauchastki protilátky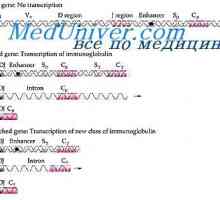 Tvorba vc-imunoglobulinových genů. Mechanismy sdružení klínové a geny protilátek
Tvorba vc-imunoglobulinových genů. Mechanismy sdružení klínové a geny protilátek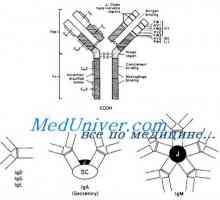 Zapletení Geny protilátky těžkého řetězce. Geny těžkých řetězců IgA
Zapletení Geny protilátky těžkého řetězce. Geny těžkých řetězců IgA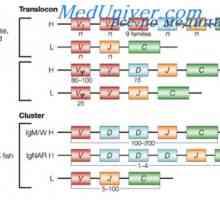 Translokon imunoglobuliny. Kombinace v- a geny protilátek
Translokon imunoglobuliny. Kombinace v- a geny protilátek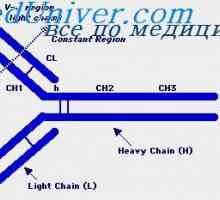 Upevňovací VH geny imunoglobulinu. Chromozom s geny protilátek
Upevňovací VH geny imunoglobulinu. Chromozom s geny protilátek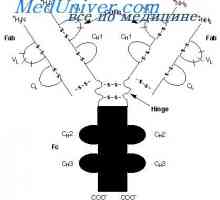 Rozdíly Variabilní oblasti protilátek. Variabilní imunoglobulinu podskupiny
Rozdíly Variabilní oblasti protilátek. Variabilní imunoglobulinu podskupiny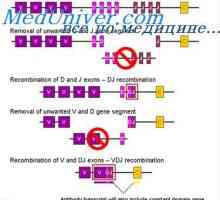 Uspořádání vc-gen. Přepínání protilátkovou aktivitu genu
Uspořádání vc-gen. Přepínání protilátkovou aktivitu genu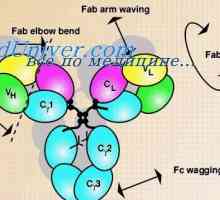 Variabilní oblasti těžkého řetězce. Variabilní oblastních genů těžkých řetězců
Variabilní oblasti těžkého řetězce. Variabilní oblastních genů těžkých řetězců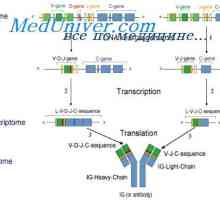 Vlastnosti protilátkou mRNA. MRNA struktura imunoglobulinů
Vlastnosti protilátkou mRNA. MRNA struktura imunoglobulinů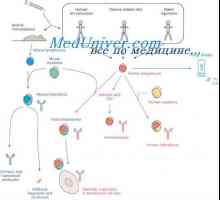 Hybridizace mRNA a DNA protilátek. Lokalizace v- a geny v genomu imunoglobulinů
Hybridizace mRNA a DNA protilátek. Lokalizace v- a geny v genomu imunoglobulinů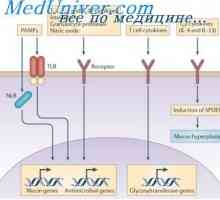 Simultánní syntéza různých imunoglobulinů. Krvetvorné kmenové buňky
Simultánní syntéza různých imunoglobulinů. Krvetvorné kmenové buňky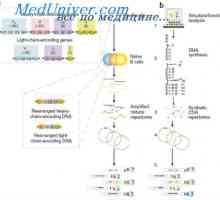 Účinek na fenotyp genové aktivity. Alelické determinanty protilátek
Účinek na fenotyp genové aktivity. Alelické determinanty protilátek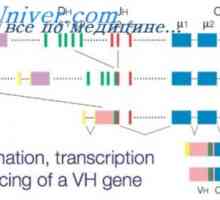 Účinek na fenotyp protilátek těžkého řetězce. Omezení V-geny
Účinek na fenotyp protilátek těžkého řetězce. Omezení V-geny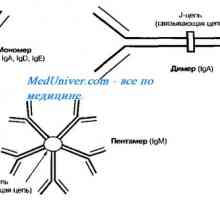 Autozomálně recesivní syndrom hyperproduction imunoglobulinu m (IgM). genová mutace pomoc
Autozomálně recesivní syndrom hyperproduction imunoglobulinu m (IgM). genová mutace pomoc Objevili nové geny zodpovědné za rakovinu prsu
Objevili nové geny zodpovědné za rakovinu prsu- Vědci našli genetickou příčinu akutní lymfoblastickou leukémií
- Vědci našli geny, které jsou zodpovědné za vzhled člověka