Ctarenie v buněčných kulturách
Video: Jeunesse Global FINITI ™ buněčného stárnutí zde končí
Použití buněčných kultur v gerontologii nabízí jedinečnou příležitost ke studiu mechanismů stárnutí a najít vlivy, které ovlivňují tento proces.Zejména se způsob buněčné kultury umožňuje řešit tyto důležité úkoly.
1. Porovnání kultury buněk odvozených od dárců různých věkových skupin, je možné rozlišovat mezi věkové změny konstitutivního indukované. Například, bylo zjištěno, že mitotická aktivita lidských fibroblastových kultur, které se za stejných podmínek, se snižuje se zvyšujícím se věkem dárcovských buněk (Schneider, Mitsui, 1976- Schneider, 1979).
V důsledku toho pozorována u zestárlých tkáních organismů pokles mitotické aktivity (Gelfant, Smith, 1972- Cameron, Thrasher, 1976) spojena s konstitutivní změny buněk samotných.
Na druhé straně, zvýšení související s věkem v indukční období tyrosinu játra pod vlivem hydrokortizonu (Adelman, 1972), by měla být považována za indukována z důvodu různého věku v kulturách hepatocytech krys pozorovány žádné rozdíly v indukci enzymu (Britton et al., 1976). To znamená, že aplikace umožňuje buněčné kultury k určení povahy stárnutí.
2. Způsob kultivace buněk poskytuje jedinečnou příležitost pro výzkum a pečlivého studia konstitutivní biochemické (Vilenchik et al., 1979) a cytologie (Schneider, 1979), změny související s věkem. Tato metoda také umožňuje přímo ověřit teorii stárnutí (Danner et al., 1978).
3. Srovnávací studie buněčných kultur získaných z různých zvířat, umožňuje identifikaci charakteristik buněk mají zvláštní význam pro biologii stárnutí. Například, bylo zjištěno, že zvířata pozitivně koreluje s prostatických fibroblastů v kultuře schopnost regenerace UV-indukovaného poškození DNA (Hart, Setlow, 1974) a v negativní korelaci s intenzitou vazbu karcinogenů na DNA fibroblastů těchto zvířat (Schwartz, Moore, 1977, 1979). Z tohoto důvodu, mechanismy na ochranu genom před poškozením jsou k gerontologie zvláštní zájem.
4. Dlouhodobá kultura (asi jeden rok) buněk in vitro lze modelovat mnoho změn probíhajících při stárnutí. I když je otázka přiměřenosti podmínek in vitro podmínek in vivo zůstává otevřenou otázkou, stárnutí buněčné kultury se zaměřuje na zvýšení počtu experimentálních, hodnocení (Hay, 1967- Beršaď, Gelfand, 1970- Cristofalo, 1972- Hayfiick, 1977- Gavrilov, Gavrilova, 1978) a teoretická práce (Olovnikov, 1973- Holliday, 1975- Hirsch, 1978- Gavrilov, Yaguzhinskii, 1978).
Kritéria a důvody pro smrt buněčných kultur
V současné době instalován Limited Střední délka života (RV) tyto buněčné kultury z normálních lidských fibroblastů (Hayfiick, Moorhead, 1961- Hayfiick, 1965- Harper, Grove, 1979), koně, několik druhů klokanů (Stanley et al, 1975), vačnatců Potorous tridactylus (Cristofalo, 1972- Stanley et al, 1975), želvy (Goldstein, 1974), mink (Hayfiick, 1975), kuřecí (Kaji, Matsuo, 1979) a myší (Hayflickův, 1975, 1977), -. epidermální keratinocyty (Rheinwald, Green, 1977), buňky gliové (Brunk`et al., 1973) a lidských jater (Kahn et al., 1977a, 1977b) normálních chondrocytů (Mayne et al., 1976) a viru Rousova sarkomu transformovaných kuřecích fibroblastů (Ponten, 1970- Leblond-Larouche, Morais , 1976).Nicméně, kritická analýza literárními údaji (Gavrilov, Gavrilova, 1978), zdá se, že stále není obecně uznávané kvantitativní kritéria pro buněčnou kulturu smrti, a samy o sobě pojmy - „smrt“ a „délka života“ kultury - vysoce podmíněné a subjektivní.
Většina výzkumných pracovníků tzv kulturu mrtvé, jestliže v libovolně nastavitelném intervalu (typicky od jednoho do čtyř týdnů) přestane zvyšovat svou sílu na požadovanou hodnotu, a to navzdory příznivým podmínkám pro růst (obvykle nastavit číslo, 2-4 krát počet počáteční kultury) ,
Podle této definice, není nutné smrt buněk v kultuře, jako jakýkoli kultura životaschopných ale nedělících se buněk, podle této definice, se považují za mrtvé. Kromě toho je kultura mrtvých pokračuje pomalu stoupat, i když mají nízkou mitotická aktivita a obsahují pouze 10 až 20% buněk schopných syntézy replikačního DNA (Cristofalo, Sharf, 1973- Milo, Hart, 1976).
Tyto kultury jsou někdy spontánně obnovit normální růst a pak provést několik zdvojení populace před další „smrti» (Holliday, Tarrant, 1972). Hmotnostní buněčná smrt na konci kultivace života, který klade velký důraz na počátku práce (Hayfiick, Moorhead, 1961- Hayfiick, 1965), byl artefakt reseeding operace.
Skutečnost, že kultivací buňky pravidelně subkultivovány, t. E. připojené ke zpracování substrátu buněčné vrstvy 0,1% roztoku trypsinu (trypsinizací) a oddělení buněk uvolnit jednoduchém k hydrodynamickým účinky (přerušení).
V prvních lisovacích velkých buněk (Milo, 1973), ve kterém se většina starých a mrtvých kultur (Bowman et al., 1975- Mitsui, Schneider, 1976). Navíc, v těchto kulturách zvyšoval počet buněk kontaktů, které nemohou být zničeny při subkultivace, aniž by se poškodily články samotné (Milo, 1973).
Z tohoto důvodu, v podmínkách pomalého růstu kultury při konstantním destrukci buněk v procesu opětovnou výsadbu dojem úplné lýze. V případech, kdy byly vyloučeny vliv reseeding operací, nebyla pozorována v rychlosti buněčné smrti z kultivačního věku žádné zvýšení.
V důsledku toho, pouze omezené slinivky buněčné kultury výhradně způsobil pokles mitotické aktivity, a žádnou ztrátu samotných buněk. Navíc existuje důvod se domnívat, že se jedná o snížení o mitotické aktivity je příčinou většiny morfologické a biochemické změny pozorované v buňkách starých a odumřelých rostlin.
Například, bylo zjištěno, že je popsáno na konci životnosti kultury „degenerativní“ buňky, která klade velký důraz na počátku studie (Hayfiick, Moorhead, 1961- Hayfiick, 1965), nezpůsobují, v důsledku úmrtí kultury, t. E. Snížení mitotický aktivitu. Bylo zjištěno, že mnohé z takzvaných „degenerativní změny“ lze získat u mladé kultury potlačena po dlouhou dobu buněčného dělení.
Všechny tyto „degenerativní změny“ zmizí po několika populačních zdvojení, pokud znovu vytvořit podmínky pro pěstování plodin (Brunk et al., 1973). Zvětšení velikosti buněk ve stárnoucí kultury také zřejmě způsobené tím, že zpomaluje a zastaví buněčné dělení. Ve skutečnosti, zvýšení velikosti byl pozorován pouze v buňkách, které nemají syntetizují DNA (Bowman et al., 1975).
Výše uvedené skutečnosti naznačují, že problém stárnutí buněčné kultury v podstatě sestoupí ke studiu příčiny, mechanismy a důsledky pozorovaného poklesu mitotické aktivity.
Tento proces nemusí nutně dojít k poškození a stárnutí buněk a může být důsledkem jejich diferenciace (Gavrilov, Gavrilova, 1978- Gavrilov, Yaguzhinskii, 1978), která je často doprovázena poklesem mitotické aktivity (Truman, 1976). Proto pojem „smrti“, „smrt“, „stárnutí“ a „naděje dožití“ buněčných kultur jsou velmi podmíněná, stejně jako pojem „mladé“ a „staré“ kultury.
Jejich použití sady výzkumníků studovat mechanismy onemocnění a úrazů, i když pouhá existence těchto úrazů a poruch v buněčné kultuře nebyla prokázána a jejich roli při omezování mitotická aktivita nebyla stanovena.
Z tohoto důvodu je vhodné použít jiné výrazy, které by se zaměřily pozornost výzkumných pracovníků je o tom, co se skutečně děje, to je. E. Na snížení mitotické aktivity. byly zavedeny tyto termíny.
Namísto toho pojmy „život“, „stárnutí“, „smrt“ a „smrti“ se navrhuje použít výrazy „replikační životnost“ buněčná kultura „replikativní stárnutí“, „smrt replikační“ a „replikační smrti“ v buněčné kultuře.
Pojem „mladé“ a „staré“ kultury je také v současné době používán s přídavným jménem „replikativní“. Young (dělení) buňky zvané replikační a staré a mrtvé (nedělící) - postreplicative.
Pro stručnost staré koncepty jsou někdy umístěny v uvozovkách, čímž zdůraznil svůj konvence (Norwood et al., 1974- Gavrilov, Gavrilova, 1978). Je významné, že i nyní biologický věk kultury je určena procenta buněk, které nejsou schopny syntézy replikačního DNA (Cristofalo, Sharf, 1973- Bowman et al., 1975).
V současné době je zatížení stanoveno následující základní projev „stárnutí“ buněčných kultur.
1. Snížené podíl buněk schopných sdílení (Merz, Ross, 1969- Cristofalo, Sharf, 1973).
2. Zvýšení doby mitotického cyklu. Radioautographic detekováno zvýšení minimální dobu generace buněk v kultivovaných lidských fibroblastů z 16 až 21 až 22 hodin (Macieira-Coelho et al., 1966). Filmové studie zjistila, zvýšení průměrné doby mezi značkami se „stárnutím“ lidských fibroblastových kultur z 16,8 až 32 hodin (Absher a kol., 1974).
3. „stárnutí“ buněčné kultury zvyšuje inhibiční účinek hustotou obyvatelstva na rychlost růstu (Macieira-Coelho et al., 1966- Absher et al., 1974- Schneider, Mitsui, 1976), takže hustota nasycení při replikaci starých kultur je přibližně 2 krát menší než "malé" kultury (Schneider, Mitsui, 1976).
Sdílet na sociálních sítích:
Podobné
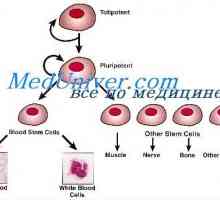 Sebeobnovy kmenových buněk. Proliferační schopnost kmenových buněk
Sebeobnovy kmenových buněk. Proliferační schopnost kmenových buněk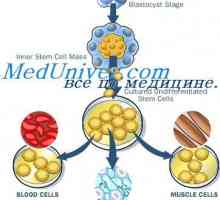 Stárnutí kmenových buněk. Mechanismy sebeobnovy kmenových buněk
Stárnutí kmenových buněk. Mechanismy sebeobnovy kmenových buněk Zahájení diferenciaci B-lymfocytů. Schéma zahájení diferenciace buněk
Zahájení diferenciaci B-lymfocytů. Schéma zahájení diferenciace buněk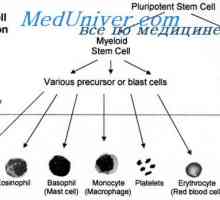 Funkce a buněk. Participatory mechanismy a buněčné imunity
Funkce a buněk. Participatory mechanismy a buněčné imunity Úloha a buňky ve zpracování antigenů. Morfologie a buněčná
Úloha a buňky ve zpracování antigenů. Morfologie a buněčná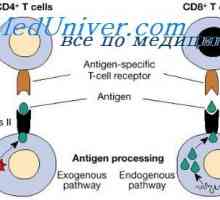 Vlastnosti spolupráce imunitní buňky. Humorální faktor protilátky
Vlastnosti spolupráce imunitní buňky. Humorální faktor protilátky Vlastnosti a charakteristiky T-supresorů. Interakce T-supresorů
Vlastnosti a charakteristiky T-supresorů. Interakce T-supresorů Experimentální přístupy k prodloužení života. neurohumorální faktory
Experimentální přístupy k prodloužení života. neurohumorální faktory- Hypotalamus-hypofýza regulace stárnutí
- Změny funkcí chromatinu v průběhu stárnutí
- Změna funkcí chromatinu v průběhu stárnutí. změna překlad
- Změna funkcí chromatinu v průběhu stárnutí. Syntéza deoxyribonukleová kyselina změny související s…
- Metabolické role v mechanismu stárnutí
- Role metabolizmu v mechanismu stárnutí. metabolismus sacharidů
 Mozek stárnutí
Mozek stárnutí- Stárnutí močového ústrojí
 Kvalitativní fenomenologické teorie stárnutí
Kvalitativní fenomenologické teorie stárnutí Stárnutí buněk v těle
Stárnutí buněk v těle- Stárnutí v buněčných kulturách. dělení Limit cell
- Účinek biologicky aktivních látek na stárnutí
- Věkové změny v genetickém aparátu buněk. změny chromozomů