Regulace specializace endoderm při tvorbě gastrointestinálního traktu u plodu
vývoj Zažívacího traktu se vyskytuje ve třech fázích. První etapa - primární specializace endoderm. Druhý krok spočívá ve vytvoření a strukturování střevní kanál, který určuje předozadní osu embrya a hranice mezi jednotlivými orgány. Třetí fáze začíná vytváření subjektů, které jsou mimo hranice střevní trubky, jako jsou játra a pankreatické vývoje.
V pokusech na laboratorních zvířatech nainstalován vývojově zachované skupiny genů, podílejících se na zaměření endoderm a několik dalších genů, podílejících se na procesu vývoje pouze obratlovců zažívacího traktu. Epitel dráhy je vytvořena z endoderm - jedna z embryonálních zárodečných listů se vyskytují při gastrulation. Studie provedené na těchto laboratorních organismech jako háďátka (půdních hlístic Caenorhabdilis elegans), octomilky (rod Drosophila), Drápatka vodní (rod Xenopus), akvarijní ryby Danio rerio a laboratorních myší, bylo možné stanovit některé základní molekulární mechanismy regulace vývojového procesu endoderm. Simulace na laboratorních myších, které se konalo knockout specifických genů pro studium jejich funkce, je považován za základ pro studium gastrointestinálních onemocnění u lidí je pylorostenóza, roztroušená atrézie, neperforované konečníku.
specializace endoderm To lze vysledovat do prvních fázích vývoje embrya. V klasických experimentálních studiích explantáty embryí kuřat ukázala schopnost než gastrulation procesu, kultura, rozvoj trávicího traktu, který ukazuje jasné určení procesu v této fázi embryonálního vývoje. Endoderm kuřecí embrya, izolovaná a umístěné v kultuře in vitro za vzniku gastrointestinálního traktu, vyznačující se expresí molekulárních markerů, které by za normálních okolností nastat in vivo, což naznačuje, že je časný jev specializace.
jeví důkaz na podporu hypotézy, že rozlišovací znaky endoderm jsou vzhledem k funkční nezávislost buňky, ale pro normální vývoj orgánů vyžaduje vzájemnou endoderm a mezoderm. V současné době je mnoho laboratorních organismy nainstalován celých skupin genů, které se podílejí na zaměření endoderm. Jedna třída genů, které kódují transkripční faktory, které se zase přímo aktivují cílových genů. Druhá třída genů kódujících syntézu signálních molekul, intercelulární spolupráce poskytuje proces. Alespoň některé z transkripčních faktorů podílejících se na procesu specializace endodermu, nadále exprimován v tkáních zažívacího traktu po celou dobu jeho následného vývoje, například genové rodiny Fox a GATA faktory (rodina transkripčních faktorů charakterizovány schopností vázat s fragmentem deoxyribonukleové kyseliny (DNA) a GATA- sekvence nukleotidů).
Regulace GIT vývoj jsou zapojeny, které působí na určitou dobu v různých lokusů, signálních drah, nebo regulačních podněty mechanismů přenosu uvnitř buněk prostřednictvím specifických molekul řetězcem mediátorů, zástupci skupiny růstových faktorů - transformující růstové faktory, b je přímo transformující růstový faktor B (TGF-b ), a kostní morfogenetický protein (BMP), stejně jako signální dráhy ježek (signalizačních systémů savců, zastoupené ve třech různých proteinů). Je prokázáno, že faktor transkripce Sox-17 hraje důležitou roli ve vývoji endoderm u myší při časných stadiích embryogeneze, ale Sox-17 role v budoucnu není nastaveno. V posledních letech se zjistilo, že signalizace Notch (transkripční faktor receptor-latentní, který je lokalizován v cytoplasmě buňky) slouží také důležitou funkci v trávicím traktu nařízení.
Až dosud nebylo možné zjistit, zda se jedná o "hlavní gen„Zahájení tvorby endoderm a následný vývoj zažívacího traktu. Ve studii zvířecích modelů některých zavedených genů, jejichž přítomnost je nezbytná a zároveň dostatečnou pro tvorbu endoderm, například mixer genu v drápy žáby. Mixl1 myší gen hraje hlavní roli v diferenciaci embryonálních kmenových buněk do endodermu. V jiných laboratorních zvířat s geny nezbytné pro tento proces, ale ne natolik, aby tvoří kompletní endoderm.
na laboratorních pokusech na zvířatech nastavit počáteční fáze indukce tvorby endodermu. Endoderm myši vytvořený z předního primitivní proužek epiblastu. Klíčovým regulátorem ranném stádiu vývoje z endoderm - uzlový-ligand superrodiny růstové faktory TGF-B. Uzlové je vyjádřena v nachází v centru blastoderm organizační nebo v uzlu primárního drážek myšího embrya. V nepřítomnosti uzlových exprese myších embryí neschopných gastrulation a tvorba difinitivnoy endoderm. Podobně Danio rerio embrya v nepřítomnosti exprese homologů uzlových také vytvořen endodermu. Na rozdíl od toho exprese funkčně aktivních receptorů TGF-b podporuje transformaci embryonálních endodermu buněk. Realizace signální dráhy TGF dosaženo stabilní meziprodukty komplexní proteiny Smad.
nezbytné pro Vývoj endoderm Je Smad2 protein a posílá tato bílkovina může vést k narušení tvorby endoderm z hindgut a různé mutace v genu, který kóduje Smad2, způsobit různé anomálie rakoviny přední / hlavy. FoxA2 také hraje důležitou roli v rozvoji endoderm, které jsou cílem signální dráhy, která vede přes uzlový faktoru Smad2 proteinu. Smad2 a Smad3 společně regulují vývoj endoderm u myší, které poskytují různý vliv na vytvoření předního střeva a zadního střeva. Kromě toho, pro vývoj jater není nutná signalizace Smad. Hlavní roli v tvorbě jater hraje Hex gen, jehož exprese je nepřítomný nebo značně redukován, když je přerušeno produktu Smad proteiny.
Etapy vývoje trávicího traktu v zárodku
| stadium | Minimální doba březosti |
| gastrulation | třetí týden |
| Tvorba kishki- primární jater a slinivky břišní záložku | 4. týden |
| Růst střevní trubice | 7. týden |
| Tvorba střevních klků | 8. týden |
| primitivní gut ponořit do břišní dutiny | 10. týden |
| Dokončení organogeneze | 12. týden |
| Vzhled žaludečních parietálních buňkách, tvorba pankreatických ostrůvků, sekrece žluči, výskyt střevních enzymů | 12. týden |
| Vzhled polykacích pohybů | 16-17-tého týdne |
| funkční zralost | 36. týden |

Je prokázáno, že entodermu Je v úzkém kontaktu s mesoderm v celém zažívacím traktu. V experimentálních studiích prováděných na tkáňových kulturách bylo prokázáno, že tvorba endoderm a jeho diferenciaci do samostatných těles je výsledkem interakce mesodermu a endoderm. Především se ukázalo, že je třeba pro signalizační účinek mesoderm endoderm podle fibroblastového růstového faktoru (FGF), pro vytvoření myši předozadní osy endodermu.
ostatní členové FGF rodina a receptory pro ně jsou nezbytné pro tvorbu jater. Další rodina ze tří genů, jsou nezbytné v procesu interakce mesoderm a endoderm - genů kódujících faktory signálních drah ježka a BMP, geny box.
v půdní hlístice Caenorhabditis elegans specializace buněk, které vyvíjejí střevní epitel, potřeboval dva geny kódující transkripční faktory GATA, zatímco GATA faktorem octomilky Drosophila rodu je kódován jediným genem had, jejichž přítomnost je nezbytná pro diferenciaci intestinálního endodermu. Ve střevě obratlovců jsou vyjádřeny třemi faktory GATA skupiny. Předpokládá se, že v procesech proliferaci a diferenciaci střevních epiteliálních GATA4 faktorů, -5 a -6 mají různé funkce, ale nebyla dosud stanovena jejich funkční význam raných fázích střevního kanálu u savců. Kromě toho faktory GATA, zástupci čeledi forkhead souvisejících faktorů (kódovány geny Fox) a signální dráhy WNT / Tcf jsou důležitými regulátory endodermu.
v obratlovců důležitou roli v iniciaci tvorby endoderm hraje zástupci superrodiny TGF-B. Molekula ELF3 cytoskelet je zapojen do signálové cesty skrz TGF-P faktorů, stejně jako potřebu rozvoje endoderm. V nepřítomnosti genu kódujícího ELF3, myši zcela chybí střevní endoderm.
mnoho faktorů, transkripce, původně považovány za specifické pro tvorbu jater, a hrát roli ve vývoji střeva. Ve studii na myších embryí, bylo zjištěno, že některé z transkripčních faktorů mají charakteristické vzory exprese, což naznačuje jejich zapojení do rozvoje střeva. Například, jaterní nukleární faktor (HNF) 3b (nyní známý jako FoxA2), se ukázal jako důležitý prvek v časných stádiích diferenciace gastrointestinálního traktu a pokračuje se vyjadřují v endoderm derivátů u dospělých. Homozygotní mutanty s úplnou absenci HNF3b není schopného vytvářet primární pásu před tvorbu střevní trubice, a několik dalších struktur. HNF3b podílí na tvorbě přední a obrany, ale ne hindgut.
Identifikuje více Číslo Fox rodinné faktory, z nichž některé se vyznačují tím, převažující nebo specifické exprese ve střevech. Jeden zástupce této rodiny, Foxl1, vyjádřený v normálním střevní mesodermu a je hlavní mediátor interakce mezi epitelem a mesenchymu. Nedostatek tohoto faktoru je doprovázen abnormální proliferace epiteliálních buněk a poruchy střevní vývoje. Tento faktor působí prostřednictvím exprese proteoglykanu vykazující se jako ko-receptory pro komponenty signální dráhy WNT, a tak komplexní aktivuje WNT / P-katenin reguluje buněčnou proliferaci. Skupina výzkumníků vedených Zaret navrhla model, ve kterém FoxA2 a GATA faktory odhalit DNA v buňkách endoderm, zajistit, že je k dispozici pro následnou vazbu transkripčních faktorů, které regulují geny buněčně specifické. To znamená, že pravděpodobný vývoj střeva četnými zástupci čeledi Fox faktory hrají důležitou roli v tomto složitém procesu, který není zcela objasněn.
U myší, některé geny, homologní gen Drosophila ocasní, vyjádřený výlučně ve střevě. CDX-1, je exprimován ve střevě dospělého organismu, je také široce exprimován v tkáních vyvíjejícího se embrya. Druhý gen, CDX-2 je exprimován v viscerální endoderm v časných stadiích embryonálního vývoje, ale následně jeho Vzor exprese je omezena výhradně na střevech. Zvýšená exprese CDX-2 vede k další diferenciaci populací buněk střeva, které se obvykle musí zůstat nediferencované. Ve stejné době, ektopická exprese CDX-2 v žaludeční stěně tkáně vede ke vzniku analogického střeva. CDX-2 je důležitým faktorem diferenciace střeva, ale až dosud není jisté, jeho role v tvorbě střevní trubky v časných stadiích embryonálního vývoje.

 Zárodečné vrstvy. Tvorba endoderm a mezodermu
Zárodečné vrstvy. Tvorba endoderm a mezodermu Význam zárodečné vrstvy. Funkce zárodečné vrstvy embrya
Význam zárodečné vrstvy. Funkce zárodečné vrstvy embrya Separace vnutrizarodyshevoy střevo. Výchova střevo v zárodku
Separace vnutrizarodyshevoy střevo. Výchova střevo v zárodku Slinných žláz embrya. Druhy slinných žláz plodu
Slinných žláz embrya. Druhy slinných žláz plodu Blastogeneze etapy. Vlastnosti proudit blastogeneze
Blastogeneze etapy. Vlastnosti proudit blastogeneze Gastrulation. Fáze gastrulace a formování orgán gastrula
Gastrulation. Fáze gastrulace a formování orgán gastrula Tvoření blastopore. Struktura a struktura blastopore
Tvoření blastopore. Struktura a struktura blastopore Mícha embrya. Diferenciace mesodermálních desek
Mícha embrya. Diferenciace mesodermálních desek Gastrulation obojživelníků. gastrulation stupně zygoty obojživelníky
Gastrulation obojživelníků. gastrulation stupně zygoty obojživelníky Chorion a amnion. Bird embryo žloutkový váček
Chorion a amnion. Bird embryo žloutkový váček Blastulation zygoty ptáky. gastrulation etapy ptáků
Blastulation zygoty ptáky. gastrulation etapy ptáků- Kapsa Amnioembrionalny. Lidské embryonální zárodečné vrstvy
 Lieberkühn kanálek. Devyatnadtsatidnevny embryo
Lieberkühn kanálek. Devyatnadtsatidnevny embryo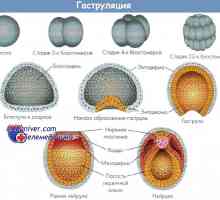 Gastrulation lidského embrya. Druhý a třetí týden vývoje
Gastrulation lidského embrya. Druhý a třetí týden vývoje Embryonální štít lidských embryí. Allantois a amnion fetus
Embryonální štít lidských embryí. Allantois a amnion fetus Vitellointestinal potrubí embryo. Lidské embryonální žloutkový váček
Vitellointestinal potrubí embryo. Lidské embryonální žloutkový váček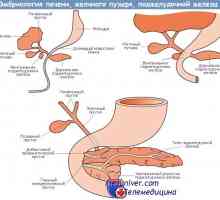 Tvorba fetální játra embryogeneze, morfogeneze
Tvorba fetální játra embryogeneze, morfogeneze- Tvorba slinivky fetálního embryogeneze, morfogeneze
- Genetická regulace tvorby lidské pankreatické
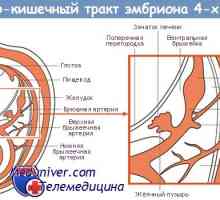 Genetická regulace tvorby gastrointestinálního traktu
Genetická regulace tvorby gastrointestinálního traktu- Genetická regulace tvorby střevní trubice