Význam Wnt signalizace v regulaci střevních kmenových buněk
Zohlednění molekulární regulace kmenových buněk a následné rozdělení buněčných populací by měl začít s procesem genu knock-TCF-4 jako důležitý mediátor Wnt signalizace ve střevě. WNT systém sekreční proteiny jsou glykoproteiny podílející se na diferenciaci a organogeneze četných tkáních. Myši s Tcf-4 genu knockout prokáže nepřítomnost proliferujících epiteliálních buněk v pozdních stádiích vývoje plodu.
proto, TCF-4 gen To je hlavní regulátor funkce kmenových buněk. Po navázání buněk s Wnt proteiny uvolňované z intracelulární komplex b-katenin se pro protein podílející se střevní adenomatózní polypózy (APC), která aktivuje Tcf-4 gen a vede k buněčné proliferaci. Při absenci zpětné vazby od Wnt genu Tcf-4 působí jako inhibitor buněčné proliferace a diferenciace buněk dochází. Dvě skupiny výzkumných pracovníků studovala inhibice Wnt signalizace inhibitor pomocí dickkopf (DKK), blokuje receptory na povrchu buněk, nebo v transgenních myších, nebo s použitím virového vektoru.
pokud Wnt signalizace blokoval působení DKK, šíření byl zastaven a krypta zmizel. Při odstranění inhibitoru došlo k zotavení a regeneraci proliferativních procesů krypt. Tyto údaje naznačují, že Wnt signalizace reguluje buněčné proliferace procesy však není nezbytné pro zachování životaschopnosti kmenových buněk v době svého vzniku. V kontrastu, odstranění proteinů APC v laboratorních myší, které se obvykle podporuje vazbu b-katenin v cytoplasmě vedla k akumulaci proteinu v buněčném jádře a aktivaci Wnt signalizace v celém epitelu.
vše epitelové buňky, se nachází v kryptách a klků na povrchu, byl transformován do proliferující nediferencovaných buněk. Kromě toho je ztráta APC proteinu v epitelu a mesenchymu narušen expresi složky extracelulární matrice. V nedávno publikované studii, kde se protein APC inaktivace dosaženo jiným způsobem, Také bylo zjištěno, že příliš rozšířená zóna proliferace v důsledku proliferace buněk diferencovaných klků. Kromě toho, že se zvyšuje počet Panethovy buněk v důsledku zvýšené exprese genů řízených signálem WNT. Stejnou funkci signalizace Wnt dráha označená skupina výzkumníků vedená van Es.

tito výzkum ukazují, že Wnt signalizace samostatně řídí program proliferace kmenových buněk a zrání programových Panethovy buňky. Mechanismus tohoto procesu však není přesně stanovena. Ve stejné době, k dispozici vědecké důkazy o tom, že pohár buňky a panethova buňka jsou odvozeny z jediné buněčné linie.
přímo epitel kryptách proteiny sekretovány WNT-6 a WNT-9b. Komplexní analýza expresního profilu messenger RNA (mRNA) umožnilo určit, zda došlo v epitelu krypt receptoru Frizzled-5 a Frizzled-6 syntézu současně v okolním antagonisty receptoru mezenchym - sFRP-1. Buňky v bazální desky klky expresi mRNA pro syntézu nezbytné WNT-2b, WNT-4 a WNT-5a. Clevers a kol. Navrhli jsme model, kde krypty epitel má určité přechodové Wnt faktory. Jejich vysoký obsah, je známo, že stimuluje proliferaci buněk. Jako buněčné migrace ve směru od základny Wnt-faktory úrovni krypty snížené, což vede k inhibici proliferace buněk a stimuluje diferenciační procesy.
Výsledky získané v minulosti výsledky výzkumu srovnatelné se zásadami tohoto modelu, kde gradienty WNT proteiny, receptory pro složky Wnt signalizace, antagonisty a-WNT provozované kmenových buněk tenkého střeva a procesy proliferaci a diferenciaci epitelových buněk.
Kromě toho, četné buněčné proteiny, role, který dosud nebyl objasněn se podílí na činnosti Wnt signalizace. EphB2 a EphVZ pod vlivem inhibičních vlastností proteinu Tcf-4 vykazují a bylo prokázáno, jsou rozhodující součásti v regulaci migrace buněk epitelu. Ef a Ephrin proteiny jsou transmembránové signalizaci proteiny vyžadované pro adhezi buňka-buňka procesů a třídění buněk. U myší, nepřítomnost produktu jako EphB2 a EphVZ Panethovy buňky migrují směrem k vrcholu v NAP větší míře, než ve směru krypty základny.
V souladu s tím knockout b-katenin, regulační protein EphVZ, To vede k tomu, že buňky jsou uspořádány Panethovy anomálním způsobem. Tato data ukazují, že zrání Panethovy buňky je autonomní buněčný proces, ne výsledek třídění a distribuce střevních krypt buněk, ve kterých řídící signalizaci dráhy Ef / Ephrin. Pravděpodobně, postižení signalizační mechanismy nejsou doprovázeny narušení procesu diferenciace buněk a tvorby klků jako celku během vývoje plodu. Předpokládá se, že tento proces je řízen jinými mechanismy.

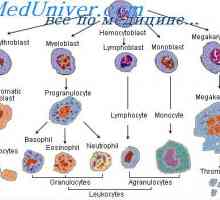 Diferenciaci kmenových buněk. Diferenciace buněk granulocytární
Diferenciaci kmenových buněk. Diferenciace buněk granulocytární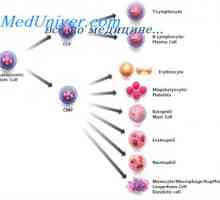 Regulace proliferace kmenových buněk. Vlastnosti kmenových buněk
Regulace proliferace kmenových buněk. Vlastnosti kmenových buněk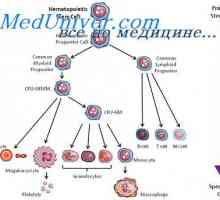 Ovládání diferenciaci kmenových buněk. Zrychlení proliferace kmenových buněk
Ovládání diferenciaci kmenových buněk. Zrychlení proliferace kmenových buněk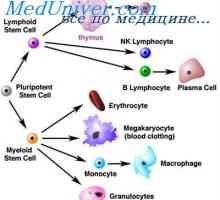 Buňky, které ovlivňují kmenových buněk. Generační cyklus kmenových buněk
Buňky, které ovlivňují kmenových buněk. Generační cyklus kmenových buněk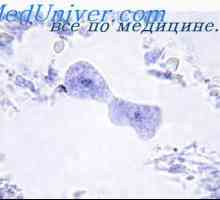 Enzymatické regulace buněčných funkcí. buněčné dělení
Enzymatické regulace buněčných funkcí. buněčné dělení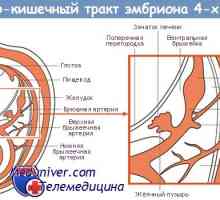 Tvorba žaludku plodu embryogeneze, morfogeneze
Tvorba žaludku plodu embryogeneze, morfogeneze Tvorba tlustého střeva v plodu embryogeneze, morfogeneze
Tvorba tlustého střeva v plodu embryogeneze, morfogeneze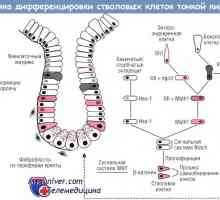 Genetická úprava diferenciaci kmenových buněk tenkého střeva
Genetická úprava diferenciaci kmenových buněk tenkého střeva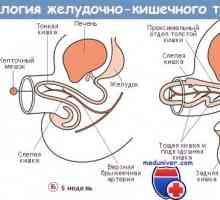 Stav střevního epitelu imunity u předčasně narozených dětí
Stav střevního epitelu imunity u předčasně narozených dětí- Mikroprostředí z kmenových buněk nice tenkého střeva
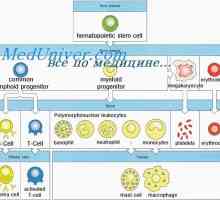 Tvorba imunity v plodu. novorozenecká lymfopoéze
Tvorba imunity v plodu. novorozenecká lymfopoéze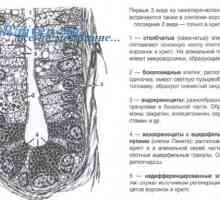 Role signální dráhy bílkoviny morfogeneze kostí (BMP), v diferenciaci kmenových buněk tenkého střeva
Role signální dráhy bílkoviny morfogeneze kostí (BMP), v diferenciaci kmenových buněk tenkého střeva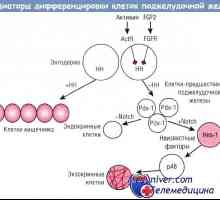 Regulace diferenciace buněk slinivky žláz s vnitřní sekrecí
Regulace diferenciace buněk slinivky žláz s vnitřní sekrecí- Regulace diferenciace exokrinních pankreatických buněk
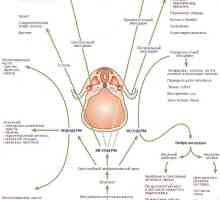 Regulace specializace endoderm při tvorbě gastrointestinálního traktu u plodu
Regulace specializace endoderm při tvorbě gastrointestinálního traktu u plodu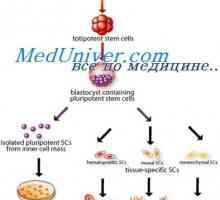 Transplantace kmenových buněk v imunodeficience a anémie Fanconiho
Transplantace kmenových buněk v imunodeficience a anémie Fanconiho Bilance dvou proteinů - klíč k boji proti leukémii?
Bilance dvou proteinů - klíč k boji proti leukémii?- Terapie embryonální kmenové buňky rozpoznán zcela bezpečné
- Kmenové buňky obnovit míchy
 Onkologiya-
Onkologiya-- Dalším krokem, který vědci pěstují endokrinních buněk