Regulační funkce přirozené imunity. Kontrola nad tvorbě adaptivní imunity
TLR signály indukují zrání dendritických buněk a expresi kostimulačních molekul nezbytných pro aktivaci T-buněk patogenspetsificheskih.
TLR-signální dráhy TIR regulované domensoderzhaschimi adaptéry, jako je MyD88, TIRAP / Mal, trif a tramvají. Diferenciace použití těchto domensoderzhaschih adaptéry TIR specificity jednotlivých TLR-zprostředkované signální dráhy. V tomto případě se extrahuje různé mechanismy, které ovládají TLR-signální dráhu a tím chrání tělo před nadměrnou aktivací přirozené imunity. Tak, TLR-zprostředkovaná aktivace vrozeného imunitního systému, reguluje nejen obranu hostitele proti patogenům, ale také na imunitní poruchy.
T-buněčné receptory rozpoznat zpracovaný antigenový peptid v komplexu s molekulou MHC II na povrchu APC. Nicméně, peptidy mohou být autologní nebo mikrobiálního původu. Vzhledem k povaze tvorby antigen-rozpoznávání repertoáru rozpoznávání T buněk peptidu v kontextu „“ autologní MHC molekuly TCR je dostatečná pro aktivaci T buněk. Pro účinnou aktivaci T-buňky se musí ještě přijmout další kostimulační signál z APC. Takový signál je CD80 a CD86 molekul. Interakce MHC II - peptidu s TCR / CD3 / CD4 komplexu poskytuje první aktivační signál onny-tého, který indukuje expresi CD40L.
CD40L interakce s CD40 na povrchu dendritických buněk dodatečně aktivuje druhý, což se projevuje na zvýšení exprese kostimulačních molekul CD80 a CD86 a IL-12 produkce. Interakce CD80 a CD86 s CD28 antigenu Th povrch poskytuje syntézu IL-2, jedna funkce, která je ke stimulaci proliferace lymfocytů a prevenci apoptózy.
posílení signály antigen a kostimulační APC je výsledkem koncentrace všech nezbytných molekul na malé ploše plazmatické membrány, známé jako lipidové plaky (lipidů) prámů. Tyto plaky obsahují vysoké koncentrace MHC ll-peptidu, HLA-DM, tetraspannye molekulami CD82 a CD86. Všechny tyto molekulárních mechanismů přispívá do koncentračních signály z APC pro aktivaci a efektorové a regulační antigen-specifických CD4 T-lymfocyty.

V současné době, známý Následující podmínky, ve kterých diferenciace dochází Th-1 buněk: 1) Aktivace velkého množství antigenního receptoru T buněk (TCR) na každé T buňce, podporovaný vysokou hustotou antigenních peptidů na povrchu dendritických buněk, a vysokým poměrem DC a T-buněk (1:10 a více), 2), výrobky z IL-12 DK, IL-23 nebo interferon I. typu.
Diferenciace T-lymfocytů v Th-2 může dojít k: 1) aktivace malého množství TCR v antigenních peptidů s nízkou hustotou na povrchu DC a nízkým podílem DC a T-buněk (1: 300 nebo méně), - 2), když je exprimován na DC OX-40L (CD25) a některé další plochy moleculesÐ 3) v přítomnosti IL-4 v době aktivace T buněk. Pravděpodobně, tvorba nepolarizované T-pomocných buněk dochází při nízké antigenpredstavleniya intenzity a v nepřítomnosti některého z třetího signálu. Pro Th-1 takových signálů jsou IL-12p70, IL-23, IFN-1-typu, IL-15, IL-18, IL-27- pro Th-2 - povrchová molekula OX-40L, které patří do rodiny TNF-a.
idea peptid v komplexu s MHC I. je třeba aktivovat CD8 CTL a MHC II-k aktivaci buněk CD4 (Th1). V současnosti se věří, že většina z antigenů se zpracovávají do pozdních endosomů a přítomné v souvislosti s de novo syntetizovaného MHC II - klasickou dráhu prezentace antigenu. Prezentace antigenů zpracovávaných počátku endosomů, provádí MHC II, recirkulace s povrchem APC.
schopnost prezentovat antigen z reentrantní cesty mají následující odborné a non-profesionální APC: nezralých DC, B buňky, monocyty, makrofágy, vsunuté epitelové buňky a epiteliální buňky thymu kůry. Existují důkazy, že časné endozomy jsou zpracovávány virové a některých bakteriálních antigenů: Některé epitopy chřipkového hemaglutininu [529], membránový protein viru spalniček povrchový protein M Streptococcus pyogenes, invazivní protein Salmonella typhimurium, sekretovaný protein Mycobacterium tuberculosis a Mycobacterium leprae, a také epitop toxinu tetanu, a další. V nedávné publikaci se začaly objevovat, což ukazuje, že zpracování a prezentace antigenu molekul MHC II může mít vliv na polarizaci odpovědi T-buněk při použití SRI recirkulační cesta prezentace odpovědi T-buněk se vyskytuje převážně typu TM, zatímco klasická dráha prezentace pomocí de novo syntetizovaného MHC II, má tendenci stimulovat reakci T-buněk podle typu Th2.
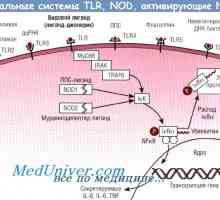 Provedení obrazraspoznayuschih PRRS receptory. Toll-like receptory
Provedení obrazraspoznayuschih PRRS receptory. Toll-like receptory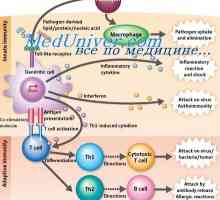 Jejich provedení Toll-like receptory. NOD-proteiny
Jejich provedení Toll-like receptory. NOD-proteiny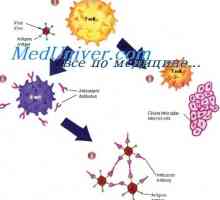 Mud88 deficitní stav. imunomodulátory
Mud88 deficitní stav. imunomodulátory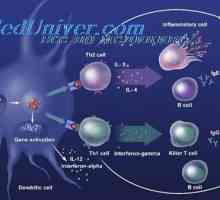 Proteiny tepelného šoku (HSP). Účinky vrozené imunitní odpovědi na HSP
Proteiny tepelného šoku (HSP). Účinky vrozené imunitní odpovědi na HSP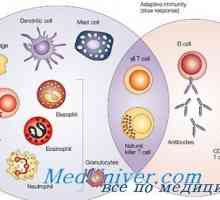 Přirozená imunita. Moderní myšlenka přirozené imunity
Přirozená imunita. Moderní myšlenka přirozené imunity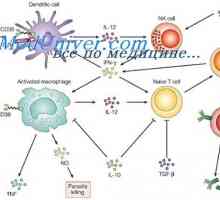 Ligandy receptoru efektorů přirozené imunity. Peptidoglykanu lipopeptidy
Ligandy receptoru efektorů přirozené imunity. Peptidoglykanu lipopeptidy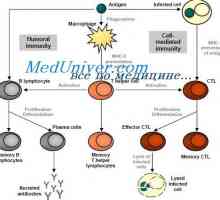 Signály Toll-like receptory. Úloha molekuly MyD88
Signály Toll-like receptory. Úloha molekuly MyD88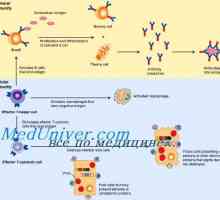 Aktivace přirozené imunity. Stupeň aktivace přirozené imunity
Aktivace přirozené imunity. Stupeň aktivace přirozené imunity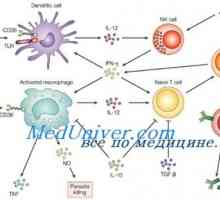 Likopid jako imunomodulátor. Mechanismy stimulace licopid imunity
Likopid jako imunomodulátor. Mechanismy stimulace licopid imunity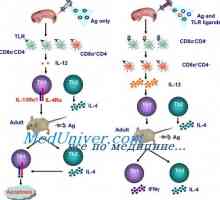 Akční imunomodulátory aktivace dendritických buněk. Zrání dendritických buněk
Akční imunomodulátory aktivace dendritických buněk. Zrání dendritických buněk Snižuje riziko alergií pomocí infekce. Imunitní odpověď na herpes simplex virus
Snižuje riziko alergií pomocí infekce. Imunitní odpověď na herpes simplex virus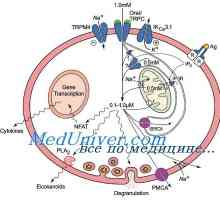 Protinádorová účinnost dendritických buněk. Účinek Immunovac-sn-4 na přirozené imunity
Protinádorová účinnost dendritických buněk. Účinek Immunovac-sn-4 na přirozené imunity Effect Immunovac-sn-4 mononukleárních buněk. Imunofenotyp leukocytů pod imunomodulátory
Effect Immunovac-sn-4 mononukleárních buněk. Imunofenotyp leukocytů pod imunomodulátory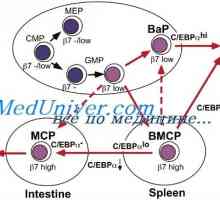 Diferenciace T a B buněk. Cytokiny indukovat diferenciaci buněk Th1 typu I-interferonu (IFN-a)
Diferenciace T a B buněk. Cytokiny indukovat diferenciaci buněk Th1 typu I-interferonu (IFN-a)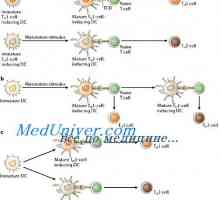 Vyhodnocení aktivaci vrozené imunity. Vyhodnocení aktivity imunitního systému,
Vyhodnocení aktivaci vrozené imunity. Vyhodnocení aktivity imunitního systému,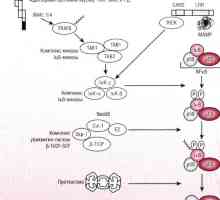 Patogeneze způsob NFKB / rel v reakci na bakterie střevního epitelu
Patogeneze způsob NFKB / rel v reakci na bakterie střevního epitelu T-lymfocyty. Charakteristika T-lymfocyty. Typy molekul na povrchu T-lymfocytů.
T-lymfocyty. Charakteristika T-lymfocyty. Typy molekul na povrchu T-lymfocytů.- Funkce v buňkách. Typy molekul na povrchu lymfocytů.
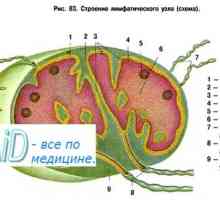 Antigen-prezentující buňky. Struktura a funkce imunitního systému.
Antigen-prezentující buňky. Struktura a funkce imunitního systému.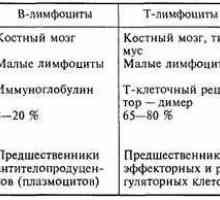 Původ (tvorba), buněk imunitního systému. Funkce buněk imunitního systému. Lymfopoéze. Bursa z…
Původ (tvorba), buněk imunitního systému. Funkce buněk imunitního systému. Lymfopoéze. Bursa z…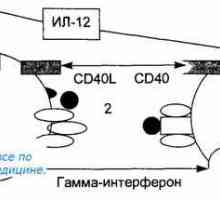 Antigen prezentace. antigen uznání. Interakce T-helper (Th1) s antigen prezentujících buněk.
Antigen prezentace. antigen uznání. Interakce T-helper (Th1) s antigen prezentujících buněk.